
Abbreviation (ISO4): Chinese Journal of Alzheimer's Disease and Related Disorders
Editor in chief: Jun WANG
Chinese Journal of Alzheimer's Disease and Related Disorders >
Astaxanthin Modulates Oxidative Stress in Primary Microglia Mediated by Lipopolysaccharide
Received date: 2020-12-25
Revised date: 2021-01-06
Online published: 2021-03-25
The increased production of Inducible Nitric Oxide Synthase (iNOS), one of the pathogenesis components of Alzheimer's disease and other neurodegenerative diseases, could be detected as one of endpoints in the oxidative-stress responses caused by the activation of primary microglia. Our aim was to explore the anti-oxidative effect of Astaxanthin (AST) on the primary microglia, derived from neonatal rats of Sprague Dawley(SD), mediated by Lipopolysaccharide (LPS).
First, primary microglia were isolated from primary mixed glia by Shake-off method, and stimulated by LPS. The primary microglia were identified through immunofluorescence while iNOS was measured by Western Blotting analysis. Secondly, this model was used to preliminarily explore the action of AST in the activated primary microglia induced by LPS.
Primary microglial cultures were confirmed using a specific CD11b antibody. iNOS was upregulated in the cells induced by LPS. The upregulation of iNOS was significantly different to that in the control (P<0.05), which suggested that the activated microglial model with oxidative stress be successfully established. Furthermore, 12.5, 25 or 50μg/ml of AST were pre-treated to the microglia for 24h before the application of LPS, which led to iNOS down-regulation at 12h, 24h or 36h, and the significant difference in the latter by comparing to that in LPS alone (P<0.05).
LPS led to the oxidative stress like iNOS upregulation in rat primary microglia. AST could down-regulated the oxidative stress such as iNOS in the microglia induced by LPS. This study suggested that AST can be with an antioxidative effect on the microglial activation in Alzheimer's disease and other neurodegenerative diseases. It may potentially be a valuable drug candidate in the clinics.
Key words: Astaxanthin; Lipopolysaccharide; Primary microglia; Oxidative stress
GUAN Xue , GUO Ling . Astaxanthin Modulates Oxidative Stress in Primary Microglia Mediated by Lipopolysaccharide[J]. Chinese Journal of Alzheimer's Disease and Related Disorders, 2021 , 4(1) : 21 -27 . DOI: 10.3969/j.issn.2096-5516.2021.01.003
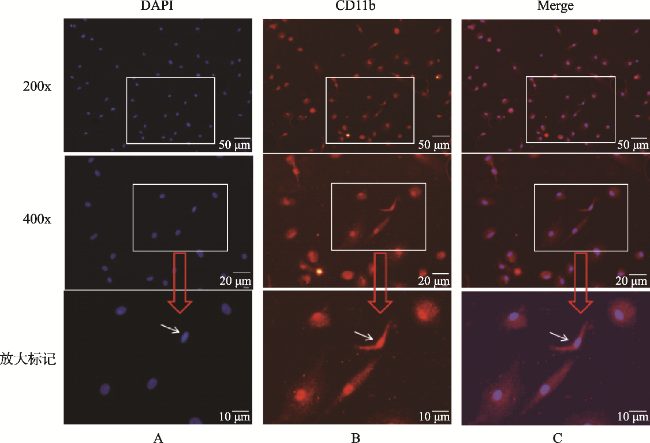
图1 原代小胶质细胞免疫荧光染色的鉴定(×200、×400)Fig.1 Identification of primary microglia cells(×200、×400) Note: Resting primary microglia derived from the cortex of neonatal rat pups showed round, fusiform or triangular shapes with bodies in red and round nuclei in blue by immunofluorescent staining. A. DAPI showed the cell nucleus in blue. B. Using CD11b antibody for immunofluorescent staining to identify the primary microglia. CD11b antibody labeled in red could combine with the protein as the antigen in the cellular fluid of the microglia so the primary microglia showed in red. C. The whole microglia were showed in red through merging |
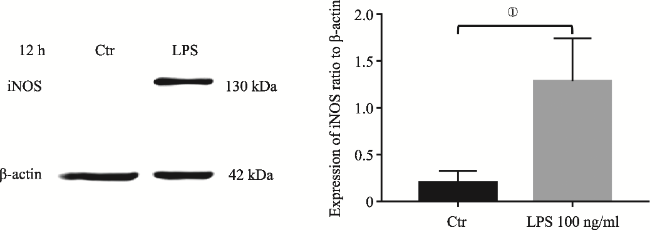
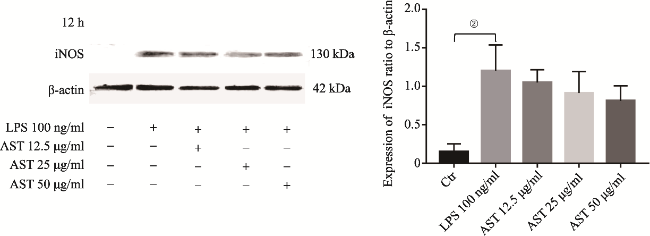
图2 LPS介导SD大鼠原代小胶质细胞12h iNOS表达Fig.2 iNOS upregulation in primary microglia induced by LPS at 12h Note: iNOS protein was measured by Western Blotting analysis. It showed iNOS upregulation in the primary microglia induced by LPS at 12h.The upregulation of iNOS was significantly different to that in the control (①P<0.05, N=3). |
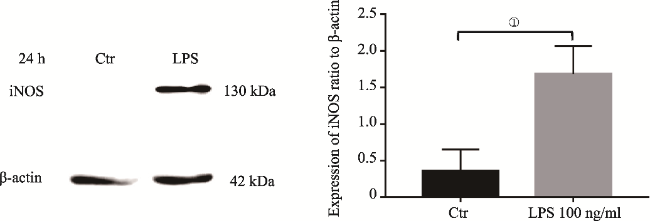
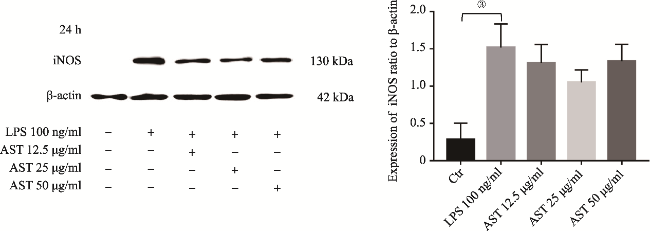
图3 LPS介导SD大鼠原代小胶质细胞24h iNOS表达Fig.3 iNOS upregulation in primary microglia induced by LPS at 24h Note: iNOS protein was measured by Western Blotting analysis. It showed iNOS upregulation in the primary microglia induced by LPS at 24h.The upregulation of iNOS was significantly different to that in the control (①P<0.05, N=3). |
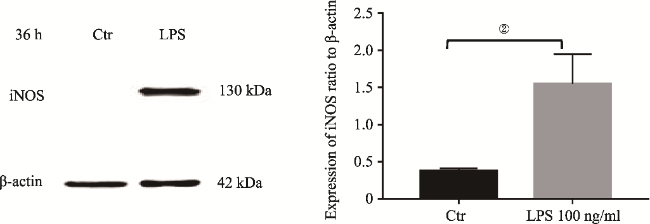
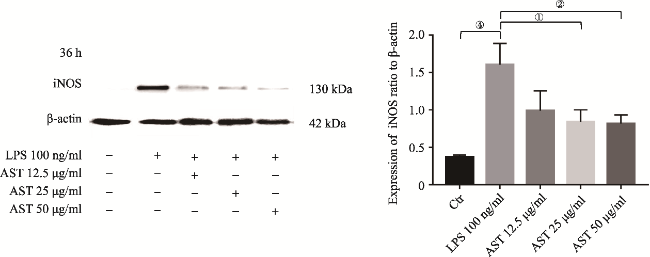
图4 LPS介导SD大鼠原代小胶质细胞36h iNOS表达Fig.4 iNOS upregulation in primary microglia induced by LPS at 36h Note: iNOS protein was measured by Western Blotting analysis. It showed iNOS upregulation in the primary microglia induced by LPS at 36h. The upregulation of iNOS was significantly different to that in the control (②P<0.025, N=3). |
图5 AST使用12h后下调LPS介导原代小胶质细胞产生的iNOSFig.5 AST down-regulating the expression of iNOS in primary microglia induced by LPS at 12h Note: 12.5, 25 or 50 μg/ml of AST were pre-treated to the microglia for 24h before the application of LPS. iNOS upregulation was showed in the cells induced by LPS alone (②P<0.025, N=4), while it was showed slight down-regulation in the cells with AST co-incubation at 12h. However, the latter did not show the significant difference by comparing to that in LPS alone (P>0.05, N=4). |
图6 AST使用24h后下调LPS介导原代小胶质细胞产生的iNOSFig.6 AST down-regulating the expression of iNOS in primary microglia induced by LPS at 24h Note: 12.5, 25 or 50μg/ml of AST were pre-treated to the microglia for 24h before the application of LPS. iNOS upregulation was showed in the cells induced by LPS alone (③P<0.01, N=4), while it was still showed slight down-regulation in the cells with AST co-incubation at 24h. However, the latter did not show the significant difference by comparing to that in LPS alone (P>0.05, N=4). |
图7 AST使用36h后下调LPS介导原代小胶质细胞产生的iNOSFig.7 AST down-regulating the expression of iNOS in primary microglia induced by LPS at 36h Note: 12.5, 25 or 50μg/ml of AST were pre-treated to the microglia for 24h before the application of LPS. iNOS upregulation was showed in the cells induced by LPS alone (④P<0.005, N=4), while it was obviously down-regulated in the cells only by 25 or 50μg/ml of AST co-incubation at 36h. The latter two showed the significant differences by comparing to that in LPS alone (respectively ①P<0.05 and ②P<0.025, N=4). Summary for |
| [1] |
|
| [2] |
应侠, 吴振, 雷严, 等. 阿尔茨海默病的发病机制及治疗药物研究进展[J]. 中国药房, 2014, 25(33): 3152-3155.
|
| [3] |
毕丹蕾, 文朗, 熊伟, 等. 阿尔茨海默病的可能药物靶点和临床治疗研究进展[J]. 中国药理学与毒理学杂志, 2015, 29(4): 507-536.
|
| [4] |
刘军. 氧化应激在阿尔茨海默病病理发生中的作用机制与干预策略[J]. 中山大学学报(医学科学版), 2020, 41(5): 661-668.
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
乔廷廷, 陈忠义, 董宝莲, 等. 依达拉奉干预LPS介导原代小胶质细胞的激活实验[J]. 昆明医科大学学报, 2018, 39(4): 5-10.
|
| [9] |
陈忠义, 乔廷廷, 董宝莲, 等. LPS介导三种啮齿动物原代胶质细胞炎症氧化反应的研究[J]. 昆明医科大学学报, 2018, 39(6): 7-13.
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
董宝莲, 郭玲. 虾青素的研究进展[J]. 中国临床药理, 2019, 35(8): 102-105.
|
| [14] |
江利华, 柳慧芳, 郝光飞, 等. 虾青素抗氧化能力研究进展[J]. 食品工业科技, 2019, 40(10): 350-354.
|
| [15] |
董宝莲, 乔廷廷, 陈忠义, 等. 建立Aβ1-42介导新生鼠原代神经元退行性变细胞模型及虾青素作用研究[J]. 阿尔茨海默病及相关病杂志, 2019, 2(2): 368-374.
|
| [16] |
|
| [17] |
|
| [18] |
廖润玲, 杨斌. 一氧化氮及诱导型一氧化氮合酶的研究进展[J]. 时珍国医国药, 2007, (4): 980-981.
|
| [19] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}