
Abbreviation (ISO4): Chinese Journal of Alzheimer's Disease and Related Disorders
Editor in chief: Jun WANG
Chinese Journal of Alzheimer's Disease and Related Disorders >
Phosphorylation of Thr231 slightly differentiates structure and interactions of cis- and trans-tau226-236 peptide hexamers
Received date: 2020-08-09
Revised date: 2020-09-03
Online published: 2020-12-25
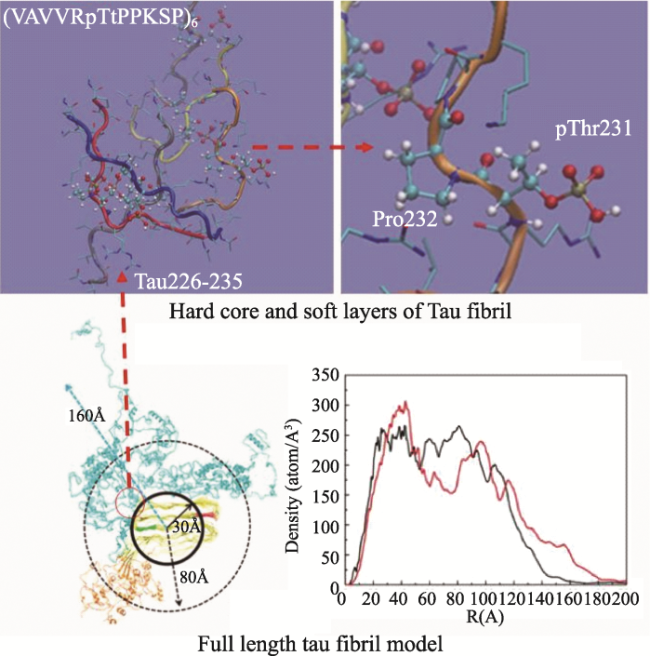
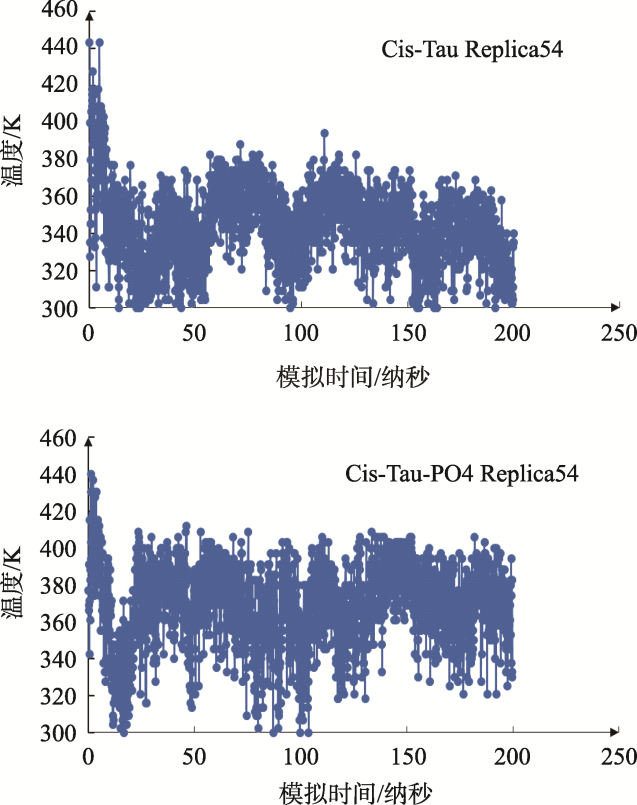
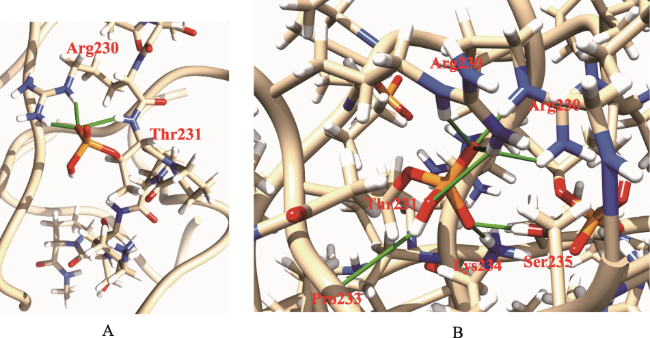
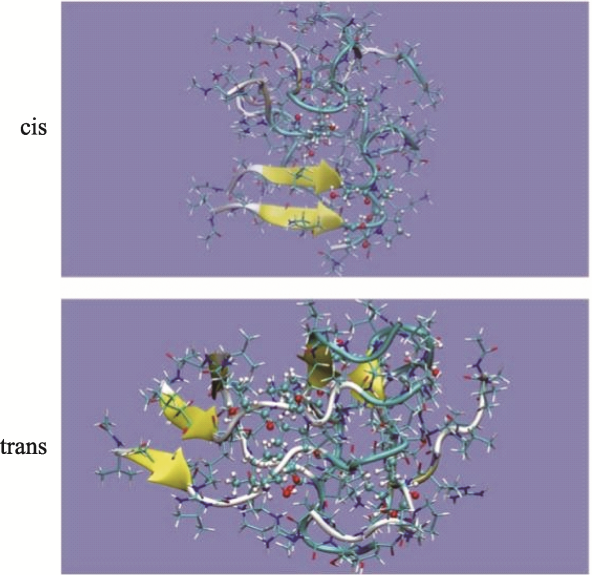
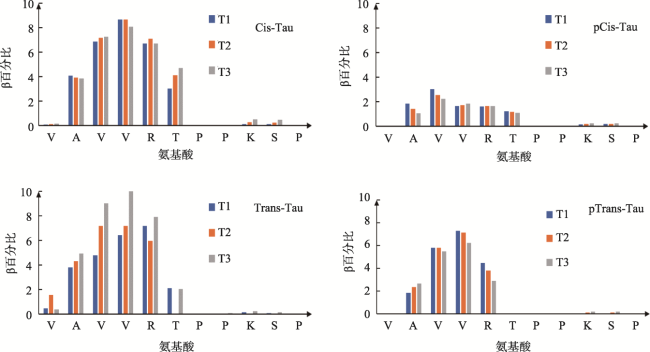
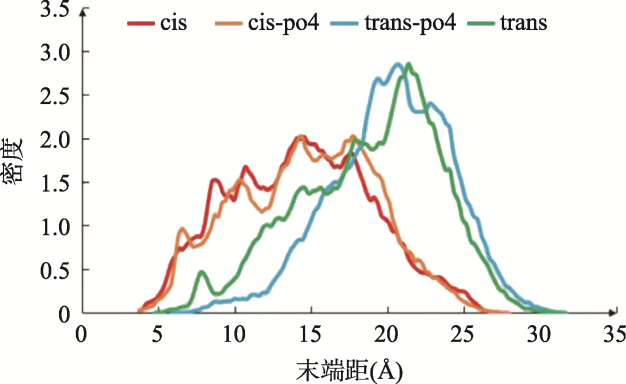
Objective: Investigate the effects of pT231 on the structure and peptide interaction in cis- and trans- tau oligomers, to examine their possible toxic mechanisms in Alzheimer's disease. Methods: Using replica-exchange molecular dynamics simulations, we studied the cis- and trans- hexamers of Tau225-246 fragment. The 56 replicas were simulated using NAMD program and CHARMM36 force field with GB model, from 300K to 450 K. Each replica was simulated for 200 ns, with total 11200 ns simulation time. Results: Interestingly, the backbone hydrogen bonds involving T231P232 had higher frequencies, indicated that they were the local interaction hot spots even before T231 phosphorylation. The backbone hydrogen bonds were more populous in trans- than in cis- tau hexamer. The 226VAVVR231T fragment had about 4%~10% β structure, more in trans- than in cis-hexamer. Although that the peptide N- to C- terminal distances were longer in trans-tau peptide, the cis- and trans-tau hexamers had similar solvent-accessible-surface areas and radius of gyrations. The Thr231phosphorylation abolished most backbone hydrogen bonds and β structures in cis-tau hexamer, but had less effects on trans-tau hexamer. The phosphate group formed salt bridge mostly with Arg230 in both cis- and trans-hexamer. However, phosphate preferd to form hydrogen bond with amide hydrogen of Arg230 in cis-hexamer, while in trans-hexamer the phosphate binds more to amide hydrogen of Lys234. The most decisive effects of T231 phosphorylation could be the exposed hydrophobic surface areaof VAVV, which increased about 100 Å2 after phosphorylation in cis-hexamer. But these areas were similar for trans-hexamer before and after T231 phosphorylation. Conclusion: Thr231 phosphorylation has different effects on the structure and chemical interaction in cis- and trans- Tau226-236hexamer. But there is no clear-cut difference that can link the local structure change to the toxicity of the tau oligomer. The differentiate effects of phosphorylation on the hydrophobic surface area of VAVV in cis- and trans- tau hexamer and the longer end-to-end distance of trans-tau226-236peptide need to be examined further in full length tau monomer and oligomers.
MENG Qingqing , ZENG Xincheng , SU Yuhong , BAI Ganggang , Nussinov Ruth , MA Buyong . Phosphorylation of Thr231 slightly differentiates structure and interactions of cis- and trans-tau226-236 peptide hexamers[J]. Chinese Journal of Alzheimer's Disease and Related Disorders, 2020 , 3(4) : 258 -266 . DOI: 10.3969/j.issn.2096-5516.2020.04.001
表1 Tau226-236六聚体磷酸化Thr231侧链基团和其他主链/侧链的氢键累计频率(%) |
| pCis-tau | pTrans-tau | ||||
|---|---|---|---|---|---|
| Donor | Acceptor | 累计频率 | Donor | Acceptor | 累计频率 |
| THR231-Main | THR231-Side | 124.6 | THR231-Main | THR231-Side | 124.2 |
| ARG230-Main | THR231-Side | 46.6 | LYS234-Main | THR231-Side | 69.9 |
| ALA227-Main | THR231-Side | 43.6 | SER235-Main | THR231-Side | 55.2 |
| SER235-Main | THR231-Side | 38.9 | ALA227-Main | THR231-Side | 52.0 |
| VAL228-Main | THR231-Side | 22.1 | ARG230-Main | THR231-Side | 29.1 |
| VAL229-Main | THR231-Side | 17.5 | VAL226-Main | THR231-Side | 28.1 |
| VAL226-Main | THR231-Side | 16.6 | VAL228-Main | THR231-Side | 10.9 |
| LYS234-Main | THR231-Side | 12.9 | VAL229-Main | THR231-Side | 5.4 |
| ARG230-Side | THR231-Side | 1015.6 | ARG230-Side | THR231-Side | 996.8 |
| LYS234-Side | THR231-Side | 209.0 | LYS234-Side | THR231-Side | 391.8 |
| SER235-Side | THR231-Side | 78.9 | SER235-Side | THR231-Side | 89.2 |
| THR231-Side | THR231-Side | 6.1 | THR231-Side | THR231-Side | 11.8 |
Note: T1: 300-304.46 K.˚ |
表2 Tau226-236六聚体多肽主链间氢键在3个温度区间的频率(%)和T231磷酸化的影响 |
| 氢键 | Cis-tau | pCis-tau | |||||
|---|---|---|---|---|---|---|---|
| Donor | Acceptor | T1 | T2 | T3 | T1 | T2 | T3 |
| SER235 | PRO232 | 25.6 | 24.2 | 23.6 | 11.7 | 12.4 | 12.3 |
| THR231 | THR231 | 24.3 | 23.2 | 17.7 | 0.2 | 0.1 | 0.1 |
| ARG230 | VAL229 | 12.3 | 11.3 | 10.1 | 2.2 | 2.6 | 2.4 |
| ARG230 | ALA227 | 11.3 | 9.8 | 7.0 | 3.4 | 2.6 | 2.0 |
| ARG230 | SER235 | 10.7 | 7.9 | 5.6 | 3.2 | 2.8 | 2.9 |
| ARG230 | VAL226 | 10.4 | 11.3 | 10.1 | 5.3 | 6.8 | 8.2 |
| VAL229 | THR231 | 9.8 | 9.0 | 8.8 | 0.2 | 0.3 | 0.6 |
| VAL228 | VAL229 | 9.7 | 8.3 | 7.1 | 1.1 | 1.0 | 1.4 |
| VAL229 | PRO233 | 8.7 | 7.8 | 6.1 | 2.4 | 2.9 | 2.6 |
| ARG230 | THR231 | 8.6 | 9.7 | 9.3 | 2.0 | 2.4 | 2.2 |
| VAL229 | VAL228 | 8.5 | 6.5 | 5.9 | 2.1 | 2.2 | 2.2 |
| THR231 | VAL228 | 8.1 | 8.1 | 7.2 | 2.2 | 2.7 | 3.1 |
| ARG230 | PRO233 | 8.0 | 6.3 | 6.3 | 5.0 | 4.7 | 3.6 |
| VAL226 | VAL228 | 7.9 | 5.9 | 3.0 | 2.4 | 3.1 | 2.1 |
| THR231 | ALA227 | 7.8 | 9.8 | 10.0 | 0.3 | 0.4 | 0.5 |
| 氢键 | Trans-tau | pTrans-tau | |||||
| THR231 | ARG230 | 23.0 | 20.3 | 15.5 | 0.0 | 0.2 | 0.4 |
| THR231 | VAL229 | 22.1 | 12.8 | 7.2 | 0.2 | 0.2 | 0.1 |
| SER235 | PRO232 | 21.9 | 18.0 | 14.8 | 7.5 | 8.7 | 8.4 |
| ARG230 | ALA227 | 17.8 | 15.0 | 12.7 | 9.2 | 9.7 | 10.0 |
| ARG230 | VAL229 | 15.7 | 17.3 | 16.2 | 0.9 | 1.0 | 0.9 |
| VAL226 | THR231 | 15.0 | 10.3 | 6.2 | 2.6 | 2.7 | 2.1 |
| ALA227 | PRO233 | 14.3 | 9.8 | 5.0 | 2.5 | 2.9 | 2.6 |
| ALA227 | VAL228 | 14.2 | 10.1 | 6.3 | 3.2 | 3.7 | 4.5 |
| ARG230 | ARG230 | 11.9 | 9.8 | 7.7 | 13.4 | 8.9 | 6.2 |
| THR231 | ALA227 | 10.5 | 9.5 | 9.5 | 0.0 | 0.0 | 0.0 |
| THR231 | PRO233 | 9.3 | 6.9 | 4.6 | 2.1 | 1.3 | 0.9 |
| VAL229 | VAL226 | 8.7 | 6.2 | 4.0 | 9.2 | 7.3 | 4.9 |
| ARG230 | VAL228 | 8.6 | 11.8 | 14.1 | 16.8 | 16.4 | 13.6 |
| VAL229 | ARG230 | 8.1 | 7.2 | 5.5 | 1.1 | 1.1 | 0.7 |
| THR231 | VAL226 | 7.6 | 8.4 | 7.6 | 1.8 | 1.4 | 0.6 |
Note: T1: 300-304.46 K°; T2: 313.57-318.23 K°; T3: 327.75-333.62 K. |
表3 Thr231磷酸化对Tau226-236多肽六聚体的结构和形态的影响 |
| alpha | beta | coil | turn | SASA(Å2)① | Rg(Å) | |
|---|---|---|---|---|---|---|
| 顺式 | 1.1 | 2.8 | 37.4 | 58.6 | 4742 (3190) | 11.5 |
| 顺式磷酸 | 0.2 | 1 | 41 | 58 | 4806 (3455) | 11.6 |
| 反式 | 2.6 | 2.4 | 50.3 | 44.7 | 4780 (3262) | 11.4 |
| 反式磷酸 | 0.2 | 1.8 | 62.4 | 35.6 | 4767 (3273) | 11.6 |
Note:①SASA Total area and hydrophobic surface area of VAVV(parenthese) |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}