
Abbreviation (ISO4): Chinese Journal of Alzheimer's Disease and Related Disorders
Editor in chief: Jun WANG
Chinese Journal of Alzheimer's Disease and Related Disorders >
Research advances on role of CD2AP in pathogenesis of Alzheimer's disease
Received date: 2020-04-07
Revised date: 2020-04-08
Online published: 2020-06-25
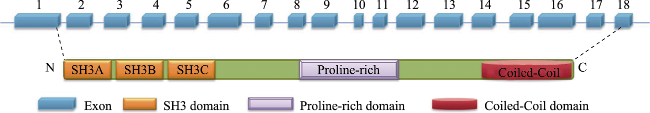
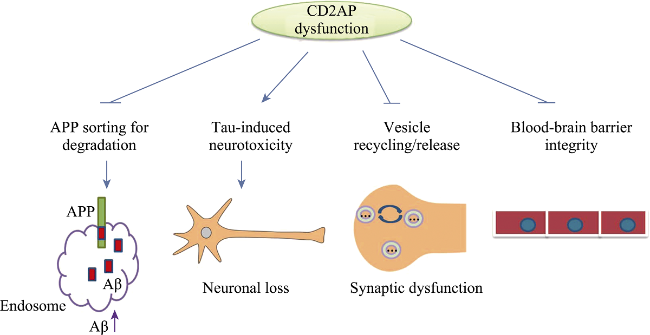
Alzheimer's disease (AD) is the most common neurodegenerative disease and has two major pathological hallmarks as senile plaques and neurofibrillary tangles that are composed of β-amyloid (Aβ) peptides and hyperphosphorylated microtubule associated protein Tau, respectively. CD2AP (CD2-associated protein) is an adaptor protein and involved in intracellular protein trafficking and cytoskeletion reorganization. Recently CD2AP has been identified as a risk factor of AD. Current evidence suggests that CD2AP may affect Aβ production and deposition and mediate Tau neurotoxicity. Moreover, CD2AP may modulate the integrity of the blood-brain barrier and the transport of synaptic vesicles. Further study on the role of CD2AP in AD may strengthen our understanding on the molecular mechanism underlying the pathogenesis of AD.
Key words: CD2AP; Alzheimer's disease; β-amyloid; Tau
ZHANG Lingliang , LUO Hong , ZHANG Xian , CAN Dan , ZHANG Yunwu . Research advances on role of CD2AP in pathogenesis of Alzheimer's disease[J]. Chinese Journal of Alzheimer's Disease and Related Disorders, 2020 , 3(2) : 141 -146 . DOI: 10.3969/j.issn.2096-5516.2020.02.010
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}