
Mercury Methylation in Periphyton and Its Impact on the Fate of Methylmercury in Aquatic Environments
Received date: 2023-10-30
Revised date: 2024-01-22
Online published: 2024-03-15
Supported by
National Natural Science Foundation of China(42277208)
National Natural Science Foundation of China(22006151)
mercury(Hg)is an important global pollutant.the aquatic environment is an important sink of mercury and the most important site for methylation and bioaccumulation.as one of the most important primary producers,periphyton is widely present in lakes,wetlands,streams,and other aquatic environments.Compared to water columns,periphyton has enhanced mercury methylation potential,which is an important source of methylmercury in aquatic environments and a key pathway for mercury entering into food chains.periphyton has diverse microbial structures and exhibits highly complex functionality.the interactions among different microorganisms result in distinct redox gradients within periphyton,forming an anoxic microenvironment conducive to mercury methylation.On the one hand,algae and bacteria in periphyton can accumulate inorganic Hg(Ⅱ)from the surrounding water,providing sufficient substrate for mercury methylation.On the other hand,periphyton is rich in metabolic secretions from various algae and bacteria,with functional groups(such As thiol groups)that can regulate the speciation of inorganic Hg(II)and enhance its bioavailability.in addition,different microorganisms can share metabolites,which can enhance the abundance and metabolic activity of Hg-methylating bacteria,thus promoting the production of methylmercury.Overall,clarifying the mercury methylation mechanism and bioaccumulation in periphyton contributes to a further understanding of the source and fate of methylmercury in aquatic environments,and provides scientific basis and data support for accurately assessing mercury pollution and environmental risks。
1 Introduction
2 Periphyton is an important site for mercury methylation in aquatic environments
2.1 Periphyton has elevated MeHg levels
2.2 Periphyton has enhanced mercury methylation potential
3 Periphyton significantly affects the fate of MeHg in aquatic environments
3.1 Periphyton is a key entrance for mercury entering into food chains
3.2 Periphyton is an important source of MeHg in water column
4 Mercury methylation in periphyton
4.1 Periphyton can accumulate Hg(Ⅱ)in aquatic environments,providing substrates for mercury methylation
4.2 Bioavailability of Hg(Ⅱ)
4.3 Activity of mercury-methylating bacteria in periphyton
5 Conclusion and outlook
Key words: mercury; methylmercury; periphyton; accumulation; methylation; food chain
Zhe Chen , Yuping Xiang , Yongguang Yin , Yanwei Liu , Lufeng Chen , Yong Liang , Dingyong Wang , Yong Cai . Mercury Methylation in Periphyton and Its Impact on the Fate of Methylmercury in Aquatic Environments[J]. Progress in Chemistry, 2024 , 36(5) : 771 -782 . DOI: 10.7536/PC231014
表1 Contents of total mercury and methylmercury in fouling organisms, water bodies and sediments in different water environmentsTable 1 Mercury and methylmercury concentrations of periphyton,water,and sediment in different aquatic environments |
| Substrates | Environments | Periphyton | Water | Sediment | Ref | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| THg/ng·g−1 | MeHg/ng·g−1 | MeHg/THg/% | THg/ng·L−1 | MeHg /ng·L−1 | THg/ng·g−1 | MeHg/ng·g−1 | ||||||||||||
| Rock | River Idrijca | 137~86100 | 2.48~495.50 | 0.18~8.76 | 6.45~63.10 | 0.03~0.10 | 5000~727000 | 3.00~10.00 | 41 | |||||||||
| Ludwigia peploides | Sanguinet lake, Aureilhan lake | 42.90~76.10a | 1.00~11.40 | 3.00~13.00 | 1.00~1.50 | 0.40~0.50 | 0.10 | 0 | 25 | |||||||||
| Teflon artificial substrates | Lake Croche | 82.80~182 | 3.61 | - | 0.84~2.26 | 0~0.77 | - | - | 49 | |||||||||
| Glass slides | The Petit-Saut | 116.00~161.00 | 32.00~64.00 | 27.59~39.75 | 0.52~2.27 | 0.12~1.57 | - | - | 50 | |||||||||
| Rock | River Idrijca | 20.00~293000 | 2.17~180.00 | - | 0~212.00 | 0.01~0.29 | 210~6380000 | 0.30~19.30 | 40 | |||||||||
| Wood and rock | Seasonal Forest Pools | 46.55~188.89 | 2.10~16.10 | 1.40~24.30 | 3.80~22.60 | 0.18~1.28 | - | - | 51 | |||||||||
| Paspalum | Tapajós River | 121~239 | 2.50~7.30 | 2.00~6.60 | 0.43~1.88 | 0.01~0.04 | - | - | 39 | |||||||||
| Sediment and wood | Little Wekiva River | 71.87~389.30 | 25~534.60 (ng·m−2) | 1.95~5.12 | - | - | - | - | 52 | |||||||||
| Sediment and wood | Santa Fe River | 3.62~12080 | 51.43~3798 (ng·m−2) | 0.96~8.07 | - | - | - | - | 52 | |||||||||
| Sediment and wood | St. Marys River | 57.74~749.30 | 35.34~218.50 (ng·m−2) | 2.92~12.03 | - | - | - | - | 52 | |||||||||
| Sediment and rock | Beaverton Creek | 308.90~739.10 | 13.63~2458 (ng·m−2) | 0.63~2.45 | - | - | - | - | 52 | |||||||||
| Sediment and rock | Lookout Creek | 28.60~425.90 | 1.33~432.30 (ng·m−2) | 0.61~3.48 | - | - | - | - | 52 | |||||||||
| Sediment and rock | Evergreen River | 117.90~510.80 | 35.23~3653 (ng·m−2) | 6.38~13.39 | - | - | - | - | 52 | |||||||||
| Sediment and rock | Oak Creek | 119.90~722.20 | 213.00~2589 (ng·m−2) | 1.33~6.73 | - | - | - | - | 52 | |||||||||
| Sediment and rock | Pike River | 92.46~261.80 | 14.73~1751 (ng·m−2) | 3.23~7.12 | - | - | - | - | 52 | |||||||||
| Teflon artificial substrates | Boreal shield lake, Lake Croche | 88.00~229.00 | 1.99~3.61 | 0.90~4.10 | - | - | - | - | 53 | |||||||||
| Rock | Boreal Shield Lakes | 42.40~271.70 | 3.10~55.30 | 2.00~36.00 | - | - | - | - | 54 | |||||||||
| Microscopy glass slides | South Carpathian region and Danube River | 54.00~262.00 | - | - | 0.44~2.19 | - | - | - | 55 | |||||||||
| Totora | High-Altitude Andean lake reservoir | 60.60~920.70 | 2.90~26.50 | 2.40~9.50 | - | - | - | - | 56 | |||||||||
| Aquatic macrophytes | Beni River, Bolivian Amazonia | 54.00~182.00 | 7~28.20 | - | - | - | 46.00~79.00 | 0.20~2.70 | 38 | |||||||||
| Paspalum. repens | Tapajós river | 67.00~198.00 | 1.00~6.00 | 6.90a | - | - | - | - | 32 | |||||||||
| Macrophyte,floating mat, soil | Everglades | 2.40~92.00 | 0.04~9.40 | - | 0.91~8.30 | 0.04~3.80 | 9.30~350 | 0.04~12.00 | 43 | |||||||||
| Rock | EFPC | - | 6.00~19.00 | - | 170.00~670.00b | 0.05~0.65 | - | - | 57 | |||||||||
| Frosted plexiglass | Near Everglades | 287.80a | - | - | 20.00a | - | 96.30a | - | 58 | |||||||||
| Rock | Streams in New Brunswick | 260.00 | 20.00 | - | - | - | - | - | 59 | |||||||||
| Creek substrate | Arivaca Lake | 110.00~1900 | 10.00~300.00 | - | - | 0.03~1.64 | 10.40~126.00 | 0.45~1.54 | 60 | |||||||||
| Cobbles | EFPC | 8000~50000b | 20.00~57.00b | - | 50.00~730.00b | 0.10~0.18b | - | - | 61 | |||||||||
| Macrophyte | Lake St. Pierre | - | 2.00~16.00 | - | - | - | - | - | 26 | |||||||||
| Rock | Eastern Canadian Shield | 15.00~398.00 | 1.00~68.00 | - | - | - | - | - | 62 | |||||||||
| Macrophyte | Lake St. Pierre | 2.00~284.00 | 0.10~24 | - | - | - | - | - | 63 | |||||||||
“-”-Not available;a-the data is average value;b-the data is estimated from The graph in The reference |
表2 Methylation ability of periphyton in different aquatic environmentsTable 2 Mercury methylation potential of periphyton in different aquatic environments |
| Substrates | Environments | Methylation incubation site (in situ or at lab) | Methylation rate constants/d−1 | Demethylation rate constants/d−1 | Net MeHg production rates/% | Ref |
|---|---|---|---|---|---|---|
| E. crassipes roots | Guapore River, Amazonia, Brazil | At lab | - | - | 6.20~25.60 | 46 |
| Paspalum. repens | Tapajós river | In situ | - | - | 0.80~22.80 | 32 |
| Ludwigia peploides | Sanguinet lake, Aureilhan lake, Escource river, France | At lab | - | 3.00~7.00 | 25 | |
| L. helminthorrhiza, P. densiflorum | Beni River Basin, Bolivian Amazon | At lab | 0.20~36.10 (% MeHg·24h−1) | - | 0.20~4.20 (sediment: 6.37~12.60) | 45 |
| E. crassipes roots | Itaúba lakes, Zé Pedro lake, Brazil | In situ | - | - | 5.10~29.20 | 64 |
| Floating mat, epiphytic periphyton | Everglades | At lab | 0~0.02 | - | 2.00~5.00 (10−3) | 65 |
| Submerged macrophytes | Lake St. Pierre (Quebec, Canada) | In situ | 15.00~18.00(10−4) | 0.08~0.22 | - | 66 |
| Totora | Lake Titicaca | At lab | 0.04~0.08 | - | - | 67 |
| Teflon artificial substrates | Boreal shield lake, Lake Croche | At lab | 0.16~5.90 (10−4·h−1) | - | 0.04~0.74 | 53 |
| Eichhornia crassipes, Polygonum densiflorum roots | Lake La Granja in Bolivia | At lab | 17.00~165.00 ( pg·g−1·12h−1) | - | - | 68 |
| Polypropylene mesh, PVC | East Fork Poplar Creek (EFPC) | At lab | 2.15~5.23(10−5) | 0.07~0.27 | - | 1 |
| E. crassipes, Myriophyllum spicatum | Lagoinha lake | At lab | - | - | 1.60~30.20 | 24 |
| Macrophytes | Lake St. Pierre | In situ | 0~0.14 | 0.10~0.14 | - | 26 |
“-”-Not available |
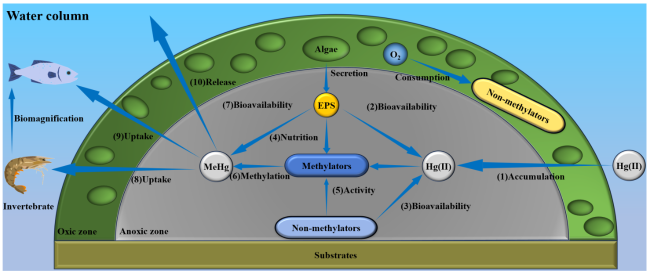
图1 水环境中附着生物的汞甲基化过程及其对水体甲基汞归趋的影响(附着生物主要通过以下途径影响水环境中的汞循环:(1) 从水体中富集Hg(Ⅱ);(2~5) 附着生物内部藻类和非汞甲基化细菌通过影响Hg(II)的赋存形态和汞甲基化细菌的活性影响汞甲基化;(6) 甲基化细菌介导的Hg(Ⅱ)甲基化;(8, 9) 无脊椎动物及鱼类对附着生物的摄食,使汞进入食物链富集累积;(10) 附着生物向水柱释放甲基汞)(根据文献[1,2]重绘)Fig. 1 Hg methylation in periphyton and its impact on the fate of methylmercury in aquatic environments (Periphyton affects Hg cycling in aquatic environments mainly through the following pathways: (1) Accumulation of Hg(II) from surrounding water. (2-5) Algae and non-methylators within periphyton affect Hg(II) speciation and the activity of Hg-methylating bacteria, thereby affecting Hg methylation. (6) Methylation of Hg(II) by Hg-methylating bacteria. (8, 9) Invertebrates and fish consume periphyton, leading to MeHg accumulation in food webs. And (10) release of MeHg into the water column) (modified from ref 1,2) |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
UNEP, Global Mercury Assessment, Geneva, Switzerland, 2019.
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
( 邹嫣, 司友斌, 颜雪, 陈艳. 环境科学, 2012, 33(9): 3247.)
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
( 张展华, 方清萱, 赵振宇, 张彤. 环境化学, 2021, 40(9): 2605.)
|
| [28] |
|
| [29] |
(董彬, 王国祥, 马杰. 淡水附植生物的组成结构特征及其生态功能. 北京: 化学工业出版社, 2018.).
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
( 程金平, 刘彩娥, 郑敏, 丁振华, 王文华. 环境科学, 2006, 27(7): 1406.)
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
( 王逢武, 刘玮, 万娟娟, 杨嘉利, 刘雪梅, 向速林, 吴永红. 环境科学, 2015, 36(11): 4043.)
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}