
Advancing Production of Sweet-Tasting Proteins Driven by Synthetic Biology
Received date: 2024-12-30
Revised date: 2025-04-06
Online published: 2025-08-08
Supported by
the National Key Research and Development Program of China(2023YFF1103700)
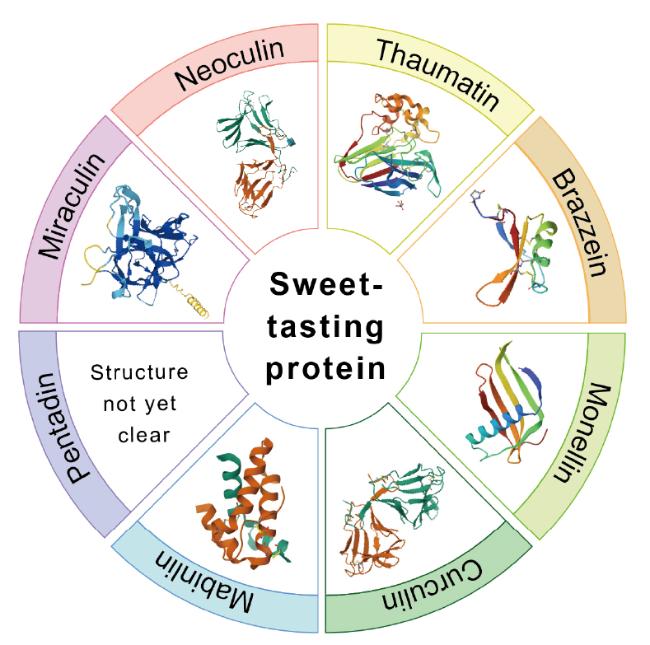
Sweet-tasting proteins,characterized by their low calorie and high sweetness attributes,demonstrate significant potential in the food industry. They not only satisfy the demand of consumers for healthy and safe sweeteners but also have the potential to replace traditional high-calorie sweeteners,thus driving innovation in the food industry. However,their commercialization process still faces challenges such as restrictions on the origin of raw materials,low yield,high extraction costs,and poor stability. In this review,the basic characteristics of sweet-tasting proteins were examined,their taste mechanisms and the relationship between their structure and sweet taste activity were investigated. Precise design and modification of sweet-tasting proteins and host through synthetic biology and artificial intelligence methods to enhance their sweetness,stability and yield were proposed. Additionally,optimizing host,expression and secretion strategies,as well as precise control of the fermentation process,can further improve the yield and activity of sweet-tasting proteins. These approaches provide a theoretical basis and technical references for addressing the existing problems in the commercial application of sweet-tasting proteins and have positive implications for promoting their widespread use in the food industry.
1 Introduction
2 Taste mechanism and structure-function relationship
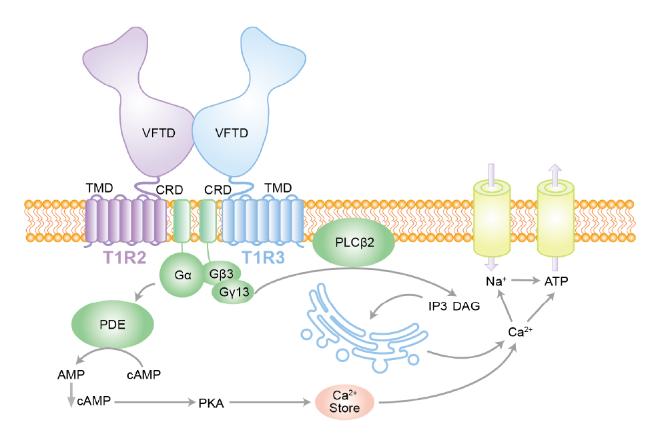
2.1 Recognition and signal transduction of sweet taste receptors
2.2 Structure-function relationship analysis
3 Customized optimization and production strategies
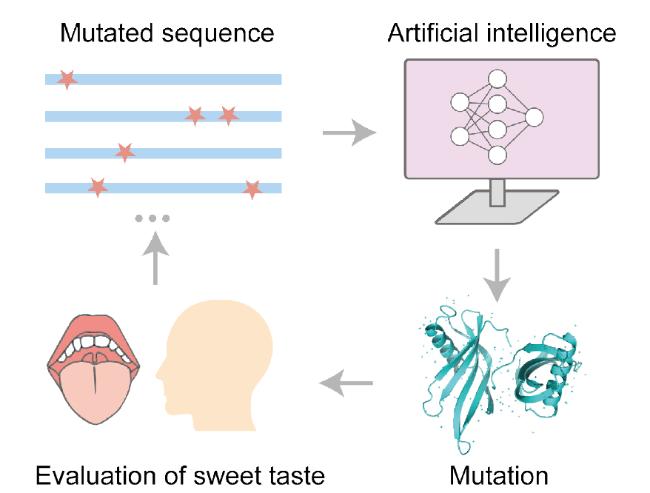
3.1 Protein precision design and modification
3.2 Optimization of strategies for host cell selection
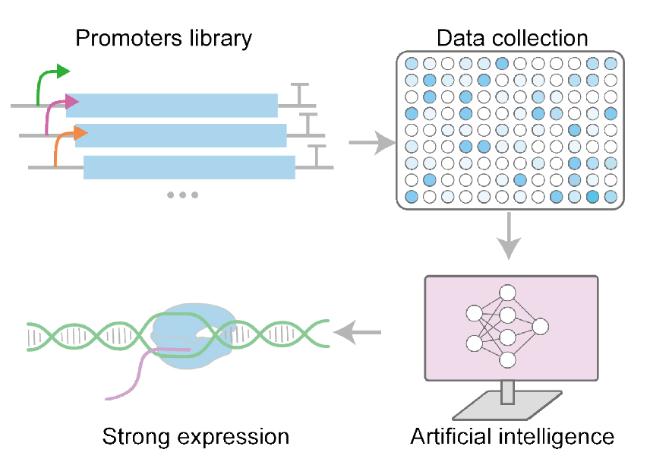
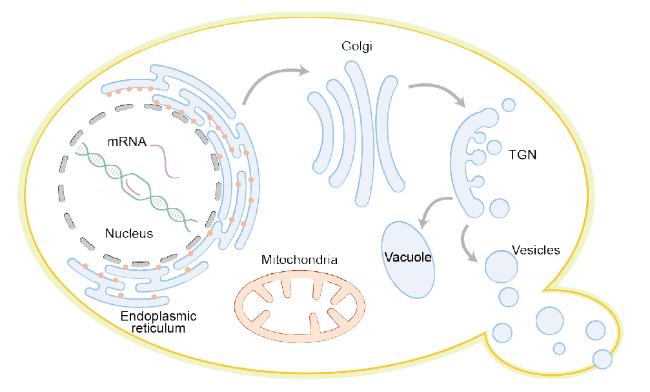
3.3 Optimization of expression and secretion strategies
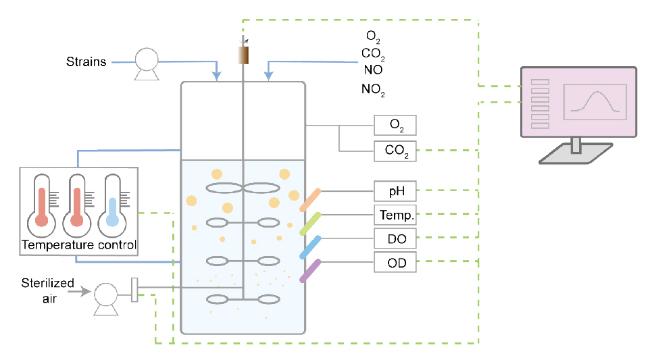
3.4 Precise control and optimization of the fermentation process
4 Conclusion and outlook
Qian Liu , Zichang Peng , Yameng Wang , Yao Geng , Xiaomin Ren , Xiaole Xia . Advancing Production of Sweet-Tasting Proteins Driven by Synthetic Biology[J]. Progress in Chemistry, 2025 , 37(8) : 1131 -1141 . DOI: 10.7536/PC241215
表1 甜味蛋白异源生产策略和产量比较Table 1 Heterologous production strategies and yield comparison of sweet-tasting proteins |
| Sweet-tasting protein | Host | Carbon source | Strategies | Yield | Ref |
|---|---|---|---|---|---|
| Thaumatin | Aspergillus awamori | Glucose and sucrose | Expression with strong fungal promoters and high gene dosage | 14 mg/L | 75 |
| Thaumatin | Aspergillus awamori | Sucrose and dextrin | Six copies of protein disulfide isomerase | 150 mg/L | 77 |
| Thaumatin Ⅰ | Pichia pastoris | Glycerol; Methanol | A co-expression strategy with the molecular chaperone,PDI | 50.7 mg/L | 100 |
| Thaumatin Ⅰ | Pichia pastoris | Glycerol; Methanol | Three copies of gene | 100 mg/L | 83 |
| Thaumatin Ⅱ | Pichia pastoris | Glycerol; Methanol | Optimal culture conditions | 62.79 mg/L | 95 |
| Thaumatin Ⅱ | Pichia pastoris | Glycerol; Methanol | Optimal culture medium selection and supplementation | 68.60 mg/L | 94 |
| Thaumatin Ⅱ | Escherichia coli | Tryptone and yeast extract | Refolding using a reduced/oxidized glutathione system | 40 mg/L | 69 |
| Brazzein | Escherichia coli | Tryptone and yeast extract | Lower induction temperature (30°C) | 15.3 mg/L | 70 |
| Brazzein | Bacillus licheniformis | Tryptone and soybean peptone | The Sec type signal peptide SPSAC guided the secretion expression | 57 mg/L | 70 |
| Brazzein | Pichia pastoris | Glycerol; Methanol | Expression using AOX1 promotor,secretion using α-factor secretion signal | 90 mg/L | 63 |
| Brazzein | Pichia pastoris | Glycerol; Methanol | Efficient secretion lysozyme using signal peptide | 345 mg/L | 84 |
| Brazzein | Kluyveromyces lactis | Glucose; Galactose | Codons optimal,expression using pKLAC2 and secretion using yeast prepropeptide secretion signal | 135 mg/L | 93 |
| Brazzein | Kluyveromyces lactis | Glycerol; Glucose and galactose | Defined medium and optimized culture conditions | 107 mg/L | 87 |
| Brazzein | Lactococcus lactis | Glucose | L. lactis NZ9000 harbouring plasmid pNZ8148 | 1.65 mg/L | 72 |
| Monellin | Pichia pastoris | Glucose | GAPDH constitutive promoter and modified N-terminus | 0.15 g/L | 107 |
| Monellin | Pichia pastoris | Glycerol; Methanol | Initiating induction at lower cells concentration and 30 ℃ | 2.71 g/L | 86 |
| Monellin | Pichia pastoris | Glycerol; Methanol and sorbitol | Fed-batch fermentation through an efficient on-line methanol/sorbitol co-feeding strategy | 2.45 g/L | 85 |
| Monellin | Saccharomyces cerevisiae | Sucrose; Galactose | Multi-copy-number integration and feed batch fermentation | 675 mg/L | 101 |
| Monellin derivative (MNEI) | Lactococcus lactis | Lactose in cheese whey | L. lactis NZ9000 harbouring plasmid pNZ8148-MNEI-ll grow on the ac-CW-based medium | 0.49 mg/L | 73 |
| Mabinlin Ⅱ | Escherichia coli | Tryptone and yeast extract | The inclusion body proteins were denatured and refolded | 59.1 mg/L | 71 |
| Mabinlin Ⅱ | Lactococcus lactis | Glucose | Optimization of inducible expression conditions | 2.72 mg/L | 71 |
| Miraculin | Escherichia coli | Tryptone and yeast extract | Improved solubilization of recombinant miraculin by the addition of arginine | 0.48 mg/L | 74 |
| Neoculin | Aspergillus oryzae | Dextrin | Constitutive induction of the unfolded protein response | 2 mg/L | 78 |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}